INTRODUCCIÓN
A
diferencia de los microorganismos, en su mayoría de vida libre como
las bacterias, los hongos y protozoos, para garantizar su
perpetuación los virus, sin excepción, requieren utilizar la
maquinaria biosintética de las células que invaden. En consecuencia,
la mayoría de los eventos en el ciclo replicativo de los virus
involucran rutas metabólicas celulares. Esto ha representado el mayor
obstáculo en el desarrollo de fármacos antivirales que inhiban
selectivamente la replicación viral, sin alterar la fisiología normal
de la célula.
Actualmente
existe una preferencia casi mayoritaria por parte de la población
mundial sobre el uso de los productos naturales en el tratamiento y
la prevención de muchas enfermedades. Prácticamente todas las
culturas del mundo han confiado históricamente en las potencialidades
de las plantas medicinales para atender necesidades primarias de
salud. Estos productos han dotado a la industria farmacéutica de
muchos de sus compuestos líderes, ya sean extraídos directamente de
las fuentes naturales o sintetizados químicamente a posteriori.1
La
selección de los productos naturales para el estudio de sus
propiedades biológicas ha sido abordada mediante 3 metodologías
fundamentales: la selección de fuentes naturales al azar, la
selección basada en quimiotaxonomía (pesquisaje de compuestos
similares en organismos pertenecientes a la misma familia o género) y
la selección basada en la etnomedicina. Este último abordaje se ha
considerado el más eficaz y consiste en el estudio de productos
naturales que tienen una larga historia de empleo en alguna comunidad
para el tratamiento de determinadas enfermedades y forman parte del
arsenal fitoterapéutico de conocimiento popular.2
El género Phyllanthus (familia Euphorbiaceae) fue descrito en 1737 por Linneo
y agrupa de 550 a 750 especies de pequeños arbustos distribuidos
ampliamente en regiones tropicales y subtropicales. El género está
dividido en 11 subgéneros, a saber Isocladus, Kirganelia, Cicca, Emblica, Conani, Gomphidium, Phyllanthodendron, Xylophylla, Botryanthus, Ericocus,y Phyllanthus. Se
estima que alrededor de 200 especies de estos subgéneros se pueden
encontrar en América, principalmente en el Caribe y Brasil. Las
infusiones de hojas, tallos y raíces de las especies de este género,
particularmente P. niruri, P. urinaria, P. corcovadensis, P. amarus, P. tenellus, P. caroliniensis, P. fraternus, P. stipulatus, y P. sellowianus,
han sido usadas durante miles de años en la medicina folklórica de
Brasil y otros países, para el tratamiento de un amplio espectro de
enfermedades que incluyen trastornos del aparato genitourinario,
infecciones intestinales, ictericia y diabetes, algunas de ellas de
posible causa viral.3
El
objetivo de esta revisión es presentar los principales resultados
encontrados en la literatura sobre las propiedades antivirales de las
preparaciones de las plantas de este género, y destacar su
demostrada efectividad sobre virus que se transmiten fundamentalmente
por la vía sexual, el virus de la hepatitis B, los virus del herpes
simple y los virus de la inmunodeficiencia humana.
MÉTODOS
Se consultaron las bases de datos PubMed (http://www.ncbi.nlm.nih.gov/entrez), FITOMED (http://www.sld.cu/fitomed) y Dr. Duke's Phytochemical and Ethnobotanical Databases (http://www.ars-grin.gov/cgi-bin/duke)
y se utilizaron como fuentes primarias de información artículos
de investigación de revistas científicas nacionales e internacionales.
ACTIVIDAD ANTI-HBV IN VITRO E IN VIVO
El
virus de la hepatitis B pertenece a la familia Hepadnaviridae, es
envuelto, con genoma de ADN de doble cadena parcial (con brechas) y
porta una ADN polimerasa ARN/ADN dependiente viriónica (reverso
transcriptasa, RT) que le permite multiplicarse a través de un
intermediario de ARN.4 Es el agente causal de la hepatitis
B, una enfermedad inflamatoria aguda del hígado que puede
evolucionar hacia la cronicidad en algunos individuos, y a largo plazo
desencadenar formas más graves de daño hepático con una elevada
mortalidad asociada: la cirrosis hepática y el carcinoma
hepatocelular primario.5,6
El estudio de la actividad antiviral de las especies del género Phyllanthus comenzó hace más de 2 décadas cuando se demostró la capacidad de un extracto de P. niruri de inactivar in vitro al antígeno de superficie del virus de la hepatitis B (AgsHB).7
Otros investigadores del mismo grupo informaron que esta
inactivación se basaba en la inhibición de la unión del AgsHB o del
antígeno de superficie del virus de la hepatitis de la marmota
(AgsHM) a sus correspondientes anticuerpos. El virus de la hepatitis
de la marmota (WHV) se ha empleado desde entonces como modelo, porque
la patogénesis de la infección en la marmota semeja considerablemente
a su contraparte humana: la hepatitis B humana. Estos autores
demostraron además que el extracto acuoso de esta planta es capaz de
inhibir la ADN polimerasa endógena de HBV y WHV. Encontraron que el
tratamiento con esta planta redujo el título viral a niveles
indetectables en el suero de las marmotas agudas o crónicamente
infectadas. Los extractos no resultaron tóxicos para el animal, los
cuales fueron aparentemente bien tolerados por la vía
intraperitoneal. Los estudios anatomopatológicos del hígado revelaron
un notorio restablecimiento de la histología normal del parénquima,
con sólo escasos y leves focos patológicos residuales, en contraste
con los daños severos observados en animales portadores no tratados.8
Posteriormente
este grupo informó que estos resultados, que fueron inicialmente
publicados atribuyéndoseles de manera errónea a P. niruri, en realidad se correspondían con la especie P. amarus.9 Los trabajos con P. amarus
continuaron y se demostró que esta planta, administrada en forma de
droga seca y molida fue capaz de eliminar el AgsHB de la sangre del
59% de portadores positivos a HBV.10,11 Estos autores publicaron un trabajo donde se recoge la actividad inhibitoria in vitro de los extractos de varias especies del género sobre la ADN polimerasa de hepadnavirus. Las especies ensayadas fueron P. debilis, P. fraternus, P. urinaria y P. niruri. Otras
especies menos conocidas (y por tanto menos usadas en la medicina
popular) también fueron evaluadas y mostraron idéntica actividad
inhibitoria sobre la enzima viral (P. mimicus y P. dontadenius).9
Con estos resultados como antecedente, se realizó un estudio comparativo del efecto inhibitorio de 5 especies de Phyllanthus endémicas de Australia sobre la ADN polimerasa del virus de la hepatitis B del pato (DHBV), P. hirtellus, P. gunnii, P. gasstroemii, P. similis y P. tenellus. Todas estas especies mostraron 50 % de inhibición de la enzima in vitro a concentraciones en el intervalo de 350-800 µg/mL, comparable a lo obtenido por Venkateswaran (1987) con extractos de P. amarus.
Los extractos fueron capaces de impedir la infección cuando se
preincubaron con los virus, al parecer mediante inactivación de la
partícula viral. Sin embargo, no tuvieron ningún efecto sobre la
infección una vez establecida, cuando se evaluó su actividad in vivo.12
En
un ensayo clínico realizado con un grupo de portadores positivos al
AgsHB a quienes se les administró diariamente durante 30 días un
extracto de P. niruri o placebo, se dejó de detectar el
antígeno en el suero de 2/3 de los portadores tratados con el
extracto, al cabo de 90 días de iniciado el tratamiento.13 A continuación otro estudio demostró que P. niruri es capaz de eliminar la hepatitis B dentro de las 3-6 semanas.14
Yeh y otros15 informaron que el extracto acuoso de P. amarus
es capaz de inhibir la producción de AgsHB pero no del antígeno E
(AgeHB) en la línea celular HepA2, que expresa y secreta
constitutivamente ambos antígenos. Estos autores plantearon que la
inhibición ocurre a nivel de la transcripción del gen viral y que
manifiesta proporcionalidad directa con el tiempo de exposición al
extracto. Sin embargo, algunos investigadores al realizar otras
experiencias con esta especie no obtuvieron iguales resultados. Milne y otros16 plantearon que la administración de extractos de P. amarus que
contenían 20 mg de geranina (unos de sus principios activos) por
dosis, 3 veces al día durante 2 meses, no tuvo ningún efecto sobre
los concentraciones séricas de AgsHB ni de AgeHB en individuos
positivos a HBV de Nueva Zelanda. Según Narendranathan,17
la administración de 3 cápsulas que contenían 300 mg de la droga
cruda en polvo, 3 veces al día tampoco redujo significativamente la
duración de la ictericia en pacientes con infección aguda.
Wang y otros14 compararon el efecto de extractos de 3 especies de Phyllanthus sobre el perfil serológico de 123 pacientes. De ellos 11 recibieron extracto de P. amarus, 42 recibieron P. niruri, 35 recibieron P. urinaria
y 35 sirvieron como controles negativos (no se les suministró
terapia). Este estudio concluyó que en la sangre de los pacientes
tratados con P. urinaria se dejó de detectar AgeHB y aumentó
la frecuencia de aparición de anticuerpos contra este antígeno
(seroconversión a anti-AgeHB), lo cual no sucedió con los pacientes
tratados con P. amarus y P. niruri. El perfil serológico con respecto a AgsHB no varió en los pacientes con ninguno de los extractos ensayados.
Por su parte Lee y otros18 y Ott y otros19 señalaron el valor de P. amarus como
antiviral al demostrar que una concentración de 1 mg/mL de un
extracto de esta planta es capaz de inhibir la secreción de AgsHB in vitro por un periodo de 48 h. Esta inhibición tiene su diana en la interacción entre los factores de transcripción a y b C/EBP y la secuencia potenciadora I (enhancer
I) del genoma de HBV, con la consecuente supresión de la
transcripción de genes virales. Este resultado concuerda con lo
planteado anteriormente por Yeh y otros (1993).
Son
evidentes las diferencias en los resultados de los diversos grupos
de investigación, sobre todo en lo concerniente a la actividad
anti-HBV in vivo de las especies de Phyllanthus: a la
capacidad de negativizar los sueros de pacientes con respecto al
AgsHB. Probablemente estas contradicciones se deban a falta de
homogeneidad en la manera de preparar los extractos, confusiones
sobre la identidad de las plantas empleadas, el tipo de suelo en el
lugar de la colecta, la época del año en que se toma la planta, la
parte de la planta utilizada, entre otros factores que influyen sobre
las concentraciones de los principios activos en la planta, así como
un escaso conocimiento sobre el periodo de ventana de acción de la
droga y la dependencia de los efectos con la concentración.
Los extractos alcohólicos de P. chamaecristoides, P. formosus y P. microdictyus, especies endémicas de Cuba, son capaces de inactivar in vitro
al AgsHB presente en sueros de pacientes positivos a HBV. Después de
ser tratados con estos extractos entre un 93 y un 97 % de estos
sueros se vuelven negativos a la detección de dicho antígeno mediante
ELISA. De estos extractos el más activo fue el de P. chamaecristoides.
En este estudio se demostró que existen diferencias significativas
(p= 0,01) en la capacidad de inactivación del AgsHB entre extractos
preparados a partir de raíces o tallos, con respecto al extracto
obtenido a partir de las hojas. Este último presentó menor actividad.
No hubo diferencias entre las actividades mostradas por los
extractos de raíces y los obtenidos a partir de tallos de la planta.20
P. orbicularis es
otra especie endémica de Cuba cuyas potencialidades en la terapia
antiviral han sido ampliamente demostradas. Los extractos acuoso y
alcohólico de esta planta también son capaces de inactivar in vitro el antígeno de superficie del virus presente en los sueros de pacientes21
y en los sobrenadantes de cultivos de células PLC/PRF/5 (células
Alexander), línea continua de hepatoma humano que expresa
constitutivamente el AgsHB.22
ACTIVIDAD ANTI-HSV IN VITRO E IN VIVO
El
virus del herpes simple (HSV) es envuelto, con genoma ADN de doble
cadena de gran tamaño, ampliamente distribuido en toda la población
mundial. Existen 2 tipos, HSV-1 y HSV-2 que son responsables de una
diversa gama de enfermedades que incluye gingivoestomatitis,
queratoconjuntivitis, encefalitis, lesiones orolabiales y genitales
entre otras. Después de la infección primaria, HSV establece una
infección persistente de tipo latente en neuronas de ganglios
nerviosos y puede reactivarse en situaciones de estrés inmunológico,
lo que provoca la enfermedad herpética recurrente. La naturaleza latente
de la infección que le permite al virus escapar de los efectores de
la respuesta inmunológica, ha imposibilitado la obtención de una
vacuna eficaz contra herpesvirus. La no disponibilidad de este
preparado, así como el desarrollo de resistencia a los antivirales
sintéticos actualmente utilizados (aciclovir, valaciclovir,
famciclovir, cidofovir) unido al alto costo de estos, que no están al
alcance de la gran mayoría pobre del planeta, han inclinado la
balanza a favor de los productos naturales.
Se ha demostrado que el extracto acuoso de P. orbicularis
inhibe la multiplicación del virus del herpes simple tipo 2 (HSV-2)
en fibroblastos de prepucio humano (FPH) y del virus del herpes
bovino tipo 1 (BHV-1) en células de riñón bovino (MDBK), con la
obtención de índices selectivos (IS) de 26,03 y 12,3 respectivamente.23 Para los estudios de actividad antiviral el IS se define como la razón entre la concentración citotóxica media (CC50) y la concentración efectiva media (CE50). El extracto acuoso de P. orbicularis fue menos activo frente a adenovirus 7 y mengovirus cultivados en células HeLa y HEp2, respectivamente.23
Este grupo también demostró que los extractos butanólico y
acético de hojas y tallos de esta especie inhiben la multiplicación
de 2 cepas del virus del herpes simple tipo 1 (HSV-1), una sensible al
aciclovir (ACV) y otra resistente al fármaco, en 2 tipos celulares
diferentes: células de riñón de mono verde africano (Vero) y FPH. Los
IS para cada fracción no variaron significativamente (p= 0,05) con
las 2 cepas de virus ni con las dos líneas celulares empleadas, según
se muestra en la tabla 1.24
{kind=link}
Por su parte Yang y otros (2005) demostraron que los extractos de P. urinaria
obtenidos con acetona, etanol y metanol empleados a una
concentración de 10 µg/mL inhibieron ligeramente (31,4; 40,4 y 41,3 %
de inhibición respectivamente) la multiplicación de HSV-1, y
resultaron muy activos (más de 90% de inhibición) contra HSV-2 en
células Vero.25
Estos
resultados ponen de manifiesto la inhibición general que sobre la
multiplicación de HSV-1 y HSV-2 tiene lugar, en presencia de
extractos de Phyllanthus. Sin embargo se han tenido que
diseñar y realizar otros ensayos para tratar de esclarecer el o los
mecanismos de acción que tributan a esta inhibición. Del Barrio y Parra (2000) encontraron que el efecto anti-HSV de P. orbicularis
es directamente proporcional a la concentración del extracto y se
hace más notorio cuando actúa sobre el virus a una baja multiplicidad
de infección (mdi= 0,1). La inhibición fue considerablemente menor
cuando se empleó una mdi= 10. En conjunto estos resultados sugieren
inmediatamente un efecto virucida directo sobre el virión en lugar de
inhibición de algún evento de la síntesis macromolecular viral. Esto
pudo ser comprobado mediante un ensayo de infectividad viral en el
cual los virus fueron preincubados con diferentes concentraciones del
extracto de P. orbicularis antes de la infección, lo cual provocó una brusca disminución en el rendimiento viral.
Idénticos
resultados fueron obtenidos para los extractos butanólico y acético
de esta planta contra HSV-1 en células Vero y FPH, que alcanzaron
elevados índices selectivos virucidas (desde 371 hasta 1040). Estos
extractos no tuvieron ningún efecto sobre la multiplicación de HSV-1
cuando se añadieron al medio 1 hora después de iniciada la infección,
confirmando su acción directa sobre la partícula viral o, en ultima
instancia, sobre algún evento temprano del ciclo replicativo
(adsorción, penetración o desnudamiento).24
La actividad inhibitoria de los extractos de acetona, etanólico, y metanólico de P. urinaria sobre
HSV-2 se verificó únicamente cuando se añadió antes o durante (0 h)
pero no 2 h después de la infección, lo cual indica un mecanismo de
acción similar al de P. orbicularis. La actividad virucida de
estos extractos (medida como reducción de la infectividad de viriones
preincubados con los extractos mediante el ensayo de formación de
placas) aumentó en proporción a la concentración utilizada y al
tiempo de preincubación pero no mostró diferencias significativas (p=
0,05) con respecto a las 2 temperaturas de preincubación utilizadas
(26 y 37 ºC). Estos resultados se resumen en la tabla 2.25
{kind=link}
En un ensayo in vivo que utilizó ratones balb/c como modelo animal se demostró la capacidad de un ungüento de P. orbicularis
de prevenir la aparición de lesiones herpéticas. En este trabajo se
les suministró de forma tópica 12 mg de ungüento/ kg de peso a cada
ratón al cabo de 8 h de haber sido inoculados mediante escarificación
de la piel y aplicación sobre esta de una suspensión que contenía 106 unidades
formadoras de placas de HSV-1. Este tratamiento se repitió durante 3
días al cabo de los cuales se comprobó una disminución significativa
(p= 0,01) de la aparición de lesiones en los ratones tratados con el
extracto respecto a los que recibieron placebo y los que no recibieron
nada. El ungüento fue capaz de evitar la aparición de lesiones en
el 60 % de los ratones tratados y un 20 % manifestó solamente lesiones
leves sin inflamación.26
ACTIVIDAD ANTI-HIV IN VITRO
El
virus de inmunodeficiencia humana tipo 1 (HIV-1) es responsable
principal de la pandemia más devastadora de los últimos tiempos: el
síndrome de inmunodeficiencia adquirida (SIDA), que desde su
emergencia ha cobrado la vida a millones de personas y actualmente
cerca de 39,5 millones viven con la infección, para la cual no existe
todavía cura definitiva ni vacuna.27
La
terapia antirretroviral grandemente activa (TARGA) ha revolucionado
la manera de tratar al paciente. Estos fármacos sintéticos han
permitido disminuir la viremia a niveles prácticamente indetectables,
reducir la transmisión (especialmente por vía vertical), aumentar el
tiempo promedio de sobrevida de los pacientes y reducir la
incidencia de infecciones oportunistas.28
Sin
embargo, otros problemas se presentan relacionados con su elevada
toxicidad y las fuertes reacciones adversas que dificultan la
adherencia de los pacientes a la terapia. Además, las evidencias de
que, aun bajo intensos regímenes de tratamiento antirretroviral, el
HIV-1 persiste, se expresa y se replica en células de memoria T CD4+
en reposo y en células mononucleares de sangre periférica, demuestran
el fracaso de estas terapias en la completa erradicación del virus.29,30
Los
elevados precios de dichos fármacos le impiden a la mayoría de los
individuos infectados acceder a ellos, de ahí que la búsqueda de
estrategias alternativas de tratamiento más seguras y al alcance de
todos sea actualmente una prioridad. Numerosos grupos se han dado a
la tarea de evaluar preparados de diferentes fuentes naturales.
Dentro de estos, los obtenidos de plantas tienen la ventaja de ser
considerablemente menos tóxicos y por tanto más tolerables, y se
puede garantizar un suministro estable del producto mediante su
cultivo.
El hallazgo de que algunas especies del género Phyllanthus
inhiben la ADN polimerasa endógena de hepadnavirus sugirió la
posibilidad de que estos extractos inhibieran también la
multiplicación de retrovirus, dado que ambos grupos de virus
comparten la necesidad de transcribir inversamente su genoma, de ARN a
ADN, como un paso esencial en su ciclo replicativo. El papel clave
de la reverso transcriptasa (RT) en la replicación de los retrovirus
ha hecho de esta enzima una diana muy atractiva en la quimioterapia de
la infección por HIV.
Esta hipótesis fue confirmada por Ogata et al (1992), quienes aislaron del extracto acuoso de P. niruri un compuesto nombrado ácido repandusínico, inicialmente obtenido de Mallotus repandus.
Este compuesto fue extraído combinando cromatografías de exclusión
con Sephadex LH-20 y celulosa, y cromatografía líquida de alta
resolución de fase reversa, y resultó 10 veces más activo frente a la
RT de HIV-1 que frente a la ADN polimerasa a de células HeLa. Las concentraciones inhibitorias medias (IC50)
obtenidas fueron 0,05 y 0,6 µM respectivamente. La inhibición fue
competitiva con respecto al complejo molde-cebador y no competitiva
en relación con el sustrato, lo cual sugiere que este compuesto se
une a la enzima en o muy cerca del sitio de unión al molde. A
concentraciones tan bajas como 2,5, 4,5 y 10,1 µM fue capaz de
inhibir en más del 90 % la producción de p24, en aproximadamente 50 %
la formación de células gigantes multinucleadas, y la aparición de
efecto citopático en células MT4, respectivamente.31
Por otra parte, el extracto acuoso de P. urinaria a una concentración de 125 µg/mL resultó activo in vitro
frente a la RT del virus de la leucemia murina de Moloney, lo que
mostró un 91% de inhibición, mientras que el extracto obtenido con
metanol no resultó activo en estas condiciones.32
Durante
un estudio en el cual se evaluó el efecto sobre la RT de HIV-1, de
extractos de 41 plantas medicinales usadas en la medicina popular
egipcia, se extrajo el compuesto putranjivain A a partir de un
extracto metanólico de frutos de P. emblica. Este compuesto inhibió específicamente la RT (IC50=
3,9 µM). La inhibición fue de tipo competitiva con respecto a
molde-cebador y no competitiva en relación con el sustrato de la
enzima (trifosfatos de desoxirribonucleótidos). Interesantemente, el
extracto de frutos de P. emblica conservó su actividad
inhibitoria después de ser incubado con albúmina de suero bovino, en
contraste con los extractos de otras especies que perdieron su
actividad debido a unión inespecífica de sus principios activos a la
proteína.33
Rev
(regulador de la expresión viral) es una fosfoproteína esencial en
la replicación de todos los lentivirus. Su función principal es
exportar al citoplasma los ARNm virales que deben sufrir solo empalme parcial o ningún empalme, secuestrándolos de la maquinaria nuclear de procesamiento postranscripcional de ARN (splicing).
Rev es capaz de reconocer y unirse específicamente a una secuencia
de ARN presente en los transcriptos virales que deben ser exportados,
denominada elemento de respuesta a Rev (ERR).34 Un compuesto aislado del extracto metanólico de hojas secas de P. niruri, nombrado nirurisida, fue capaz de inhibir específicamente la unión de Rev a la secuencia ERR in vitro (IC50=
3,3 µM), pero no de proteger cultivos de células CEM-SS de la
infección aguda por HIV, a concentraciones de hasta 260 µM.35
Probablemente los estudios más detallados sobre la actividad anti-HIV de Phyllanthus son los realizados por el grupo de Notka et al, quienes confirmaron la propiedad inhibitoria sobre la RT de HIV-1 in vitro de los extractos acuoso (IC50= 4,6 µg/mL), metanólico (IC50= 12,6 µg/mL e hidroetanólico (IC50= 8,9 µg/mL) de P. amarus.
A partir del extracto hidroetanólico purificaron 2 galotaninos que
resultaron más activos que el extracto de partida: corilagin (IC50= 5,9 µg/mL) y geranin (IC50=
1,8 µg/mL). De acuerdo con lo informado previamente para otros
compuestos presentes en las plantas del género, la inhibición de la
RT fue de tipo no competitiva con respecto al sustrato y en relación
con el complejo molde-inhibidor predominó la de tipo competitiva.36
La
capacidad de estos compuestos de inhibir la multiplicación de HIV-1
en células MAGI también fue comprobada. Todos los extractos de P. amarus
fueron muy efectivos, para un IS entre 32 y 517. Los galotaninos
purificados, corilagin y geranin, mostraron los mejores IS
antivirales: 1104 y 1250 respectivamente, lo que demuesta el carácter
específico de su actividad inhibitoria. Estos extractos fueron
también ensayados frente a varias cepas de HIV-1 y HIV-2 resistentes a
los principales inhibidores sintéticos de la RT, análogos y no
análogos de nucleósido actualmente incluidos en la TARGA, y se
obtuvieron resultados similares en cuanto a inhibición de la
multiplicación viral. El extracto obtenido con hexano no tuvo ninguna
de las actividades anteriores.36
Aunque P. amarus
bloquea eventos intracelulares de la replicación viral, como se ha
descrito hasta aquí, la inhibición más relevante se ha verificado a
nivel de la internalización viral. El extracto hidroetanólico y la
geranina inhiben la entrada del virus a la célula en más del 80 % a
sólo 10 µg/mL. A una concentración tan baja como 2,5 µg/mL, todavía
se manifiesta cerca de un 75 % de inhibición, lo cual representa una
eficacia que supera a la del heparan sulfato, compuesto empleado como
control positivo en este ensayo.36 Esta inhibición de la
penetración se debe a la unión de los galotaninos a la gp120, lo cual
impide la adsorción al receptor primario del virus (CD4). La
efectividad aumentó proporcionalmente con la pureza de los galotaninos.
Las IC50 obtenidas fueron 2,65, 1,36, 0,48 y 0,5 µg/mL
para el extracto hidroetanólico, la fracción de galotaninos, geranin
purificado y corilagin purificado, respectivamente. Mediante un
ensayo de fusión de células infectadas por HIV que expresan gp120 en
la superficie y células sin infectar CD4+, se comprobó que estos
compuestos no actúan sobre la fusión, ya que al ser añadidos después
de establecida la interacción gp120-CD4, no impiden la formación de
células gigantes multinucleadas producto de la fusión celular.37
El efecto de los extractos y compuestos de P. amarus
sobre otras enzimas de HIV-1, integrasa (IN) y proteasa (PR) también
fue estudiado in vitro. Ambas enzimas resultaron inhibidas en
diferente magnitud por todos estos compuestos, aunque IN fue más
sensible a la inhibición. El efecto inhibitorio sobre ambas proteínas
aumentó proporcionalmente con la pureza de los galotaninos (corilagin
y geranin), y el extracto hidroetanólico de partida resultó el
menos efectivo. Esto demuestra que esta clase de compuestos es
responsable de la mayor parte de la acción antiviral de estas
plantas.37
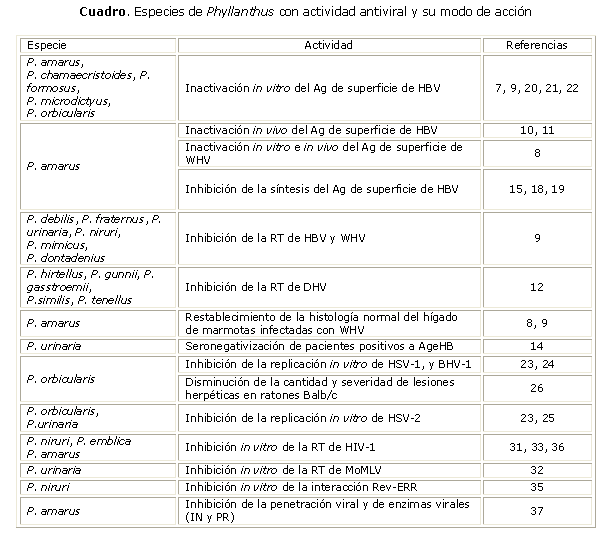
En el cuadro se resumen las principales especies de Phyllanthus que han mostrado actividad contra hepadnavirus, herpesvirus y retrovirus y sus posibles mecanismos de acción propuestos.
{kind=link}
CONCLUSIONES
La
investigación científica en el campo de los productos naturales es
en realidad un estudio retrospectivo, pues la mayor parte de los
resultados obtenidos esclarecen los mecanismos de acción y principios
activos de productos cuyo empleo por el hombre se encuentra
documentado históricamente, y en ocasiones data de los orígenes
mismos de las comunidades humanas. Los laboratorios están
desentrañando las razones para el uso, en ocasiones milenario, que se
le ha dado a cada planta en el marco del conocimiento popular
transmitido de generación en generación.
Los
datos aquí presentados, resultado del trabajo de más de 2 décadas de
numerosos grupos de investigación, demuestran de forma contundente
el valor de las plantas del género Phyllanthus en el control
de 3 patógenos que representan actualmente graves problemas de salud a
nivel mundial: el virus de la hepatitis B, los virus del herpes
simple y los virus de inmunodeficiencia humana. Cada día son más los
reportes en la literatura de variantes o cepas de patógenos
resistentes a fármacos. El mejor ejemplo es la resistencia
desarrollado por muchos aislamientos de HIV a los antirretrovirales
sintéticos empleados, que conllevó a la necesidad de combinar varios
de ellos en los llamados cocteles (multiterapia) con el consecuente
aumento del daño tóxico y reacciones adversas como la lipodistrofia y
la toxicidad mitocondrial. Los extractos de especies de Phyllanthus
han demostrado ser verdaderos «cocteles naturales» pues contienen
múltiples principios activos que inhiben diferentes moléculas virales
en diferentes momentos del ciclo replicativo, lo que hace
prácticamente imposible el desarrollo de resistencia.
Aun
queda mucho por hacer y deben realizarse estudios más detallados en
lo concerniente a la estabilidad de los productos, la formulación,
determinación de las mejores vías de presentación, la
biodisponibilidad y la farmacocinética in vivo. Sin embargo,
ya se comienza a ver la medicina natural y tradicional, no como la
alternativa más barata y únicamente asequible a los pobres, sino como
una verdadera promesa en la manera de abordar el tratamiento de las
enfermedades infectocontagiosas.

No hay comentarios:
Publicar un comentario